



2. Department of Neurology, The Second People's Hospital of Jining, Jining, China;
3. Shandong Institute of Behavioral Medical Science, Jining Medical College, Jining, China
Objective: To study the effects of regional cerebral blood flow (rCBF) perfusion on learning and memory function in special brain areas and its molecular mechanism in rat. Methods: Sixty-four adult male healthy Sprague-Dawley (SD) rats were randomly divided into two groups: A false operation group and an operation group. The false operation group was randomly divided into four subgroups (A0, B0, C0, and D0) and the operation group was randomly divided into four subgroups (A, B, C, and D), with eight rats in each subgroup. The operation group underwent bilateral common carotid artery permanent ligation, while the other group only underwent a skin incision without the bilateral common carotid artery permanent ligation. Learning memory function of rats in each subgroup was measured using a Y-maze at 4 h, 8 h, 24 h, and 3 days after surgery. The rCBF in the right frontal lobe and hippocampus was detected using the Periflux PF model laser Doppler flowmetry and c-fos, c-jun, Bcl-2, and Bax protein expression in the right frontal lobe and hippocampus was measured using immunohistochemistry. Results: The rCBF in the right frontal lobule division and right hippocampus division was significantly lower in the operation group than in the false operation group (P <0.05). The error number (EN), time to reach the target, and total reaction time (TRT) for the learning index using the Y-type labyrinth test in the operation group were significantly higher than that in the false operation group (P <0.05); however, the active avoid rate in the operation group was significantly lower than that of the false operation group. Expression of c-fos and c-jun as well as the average absorbency in the right frontal lobule division and right hippocampus division in the operation group were significantly higher than those in the false operation group (P <0.05). The number of Bax and Bcl-2-positive cells was significantly higher in the operation group, and the expression ratio of Bax/Bcl-2 in the operation group was significantly higher than that in the false operation group (P <0.01). Conclusions: rCBF decrease can impair learning and memory function in rats, which may be related to the increased expression of c-fos, c-jun, Bcl-2, and Bax proteins in the t he frontal cortex and hippocampus.
(2) Immunohistochemistry for c-fos and c-jun: The experimental steps were as follows: (a) the dewaxed slices were treated with 3% hydrogen peroxide (H2O2) for 10 min to destroy the endogenous peroxidase activity; (b) antigen retrieval was performed by boiling the slices for 10 min in EDTA in a microwave,and then,naturally cooling it to room temperature; (c) the slices were placed in natural sheep blood serum (10%) at room temperature for 30 min; (d) the slices were then incubated in rabbit anti-c-jun (1:200) and anti-c-fos (1:50) antibodies for 24 h at 4 ℃; (e) incubation with the secondary antibody consisting of 1:100 biotin according to the direction of SP reagent box was performed for 120 min at room temperature with shaking,followed by washing with PBS for 10 min each,3 times; (f) incubation with 1:100 streptavidinperoxidase was for 60 min at room temperature with shaking,followed by washing with PBS 4 times for 10 min each; (g) the slices were counterstained with hematoxylin for 1 min; (h) the slices were mounted on slides,dehydrated,and coverslipped,followed by analysis under a microscope with computer image processing system. PBS (0.01 mol/L) was applied as the negative contrast.
(3) Quantitative measurement of c-fos and c-jun positive cells: c-fos and c-jun positive cells were randomly measured in 20 units of areas on each slice under 400× magnification (units of areas are the window of areas provide by the computer system),and the average numbers of positive cells in the 20 units for c-fos and c-jun were calculated per slice. Semi-quantitative analysis of the randomly selected 20 slices from each group was done by testing the pigmentation average absorbency with the MPIAS-500 colorized multimedia image processing system.
(4) The detection of Bcl-2 and Bax: The experimental steps were as follows: The dewaxed coronary slices were treated with 3% hydrogen peroxide (H2O2) for 15 min to destroy the endogenous peroxidase activity, then washed with PBS,incubated in rabbit anti-rat Bcl-2 or Bax antibodies for 2 h at 37 ℃,and then washed with PBS. In order to eliminate nonspecific pigmentation,some of the slices were replaced with PBS and incubated in a similar manner. This was followed by incubation with instant type Ⅱ antibody for 30 min at 37 ℃,washing with PBS,staining with 3,3’ diamino benzidine (DAB),dehydration,and coverslipping. Randomly,10 slices from each rat were used to count the number of Bcl-2 or Bax positive cells in 100 cells in each hippocampus CA1 region of the same visual field under the microscope.
The positive cells were identified by the brown yellow granules in the cytoplasm or on the karyotheca under the optical microscope.
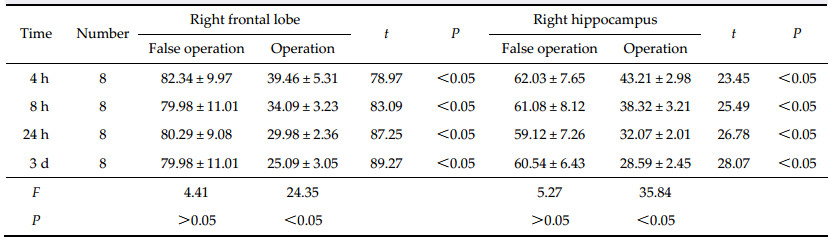
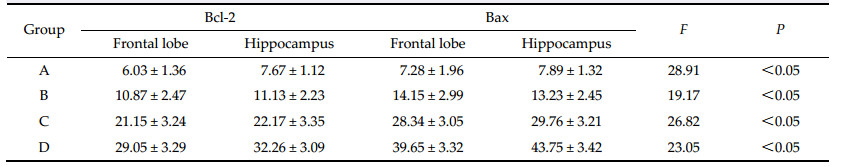
2.3 Statistical analysis The results are shown as mean ± standard deviation. The students t-test was used to compare the differences between two groups,and F-analysis was used to compare the mean between multiple groups. Analysis was performed with SPSS 12.0 software. 3 Results 3.1 The effect of the decreased rCBF in the right frontal lobe and hippocampus on learning and memory function Learning and memory performance of rats in the A,B, C,and D groups was lower than that of rats in the A0, B0,C0 and D0 groups at 4 h,8 h,24 h,and 3 d after operation. The indexes of training times,EN,time to reach the mark,and TRT also increased gradually in the A,B,C,and D groups,and the initiative evasion rate (IER) decreased gradually (trend test P < 0.05). However,there were no significant differences in learning and memory indexes among the A0,B0,C0, and D0 groups (Table 1).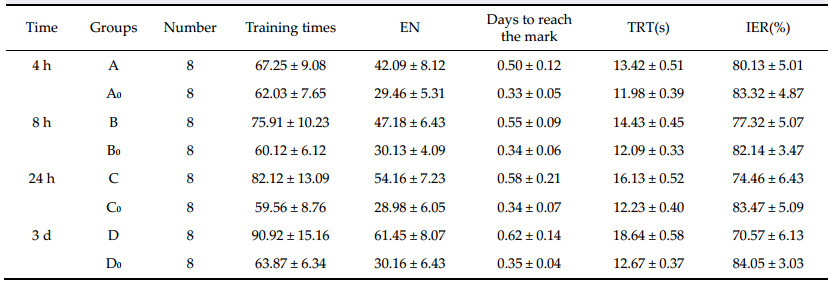 |
 |
 |
 |

|
| Figure 1(a–b) c-fos protein positive cells in the hippocampus tissue of A0 and A subgroups (serosity SP dyed ×400). |

|
| Figure 2(a–b) c-jun protein positive cells in the hippocampus tissue of the A0 and A subgroups ( karyon SP dyed ×400) |
 |

|
| Figure 3(a–b) Bcl-2 expression in the immunohistochemical slice from the hippocampus cell serosity (SP ×400). |

|
| Figure 4(a–b) Bax expression in the immunohistochemical slice from the hippocampus cell serosity (SP ×400). |
 |
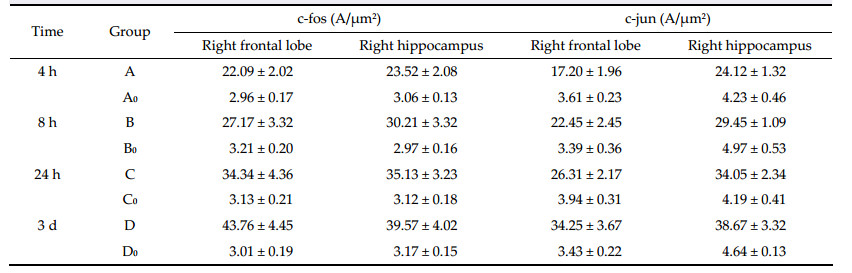
There are usually two types of ischemic brain tissue injuries: necrosis and apoptosis. There is significant apoptosis-related protein expression during cerebral ischemia,which supports the occurrence of apoptosis in ischemic trauma. However,the mechanism of neuronal apoptosis after cerebral ischemia remains unknown. It has been demonstrated that the active expression of apoptosis-related genes plays a critical role in the mechanism of neuron apoptosis. There are usually two types of apoptosis related genes: the apoptosis promoting genes like c-fos,c-jun,c-myc, p53,bax,and fas; the anti-apoptosis genes,such as Bcl-2 and Bcl-x1[5]. c-fos and c-jun can be expressed transiently when the cell is stimulated by a set of physical,chemical,and biological factors,to produce c-fos and c-Jun protein[6, 7, 8]; therefore,the levels of c-fos and c-jun expression are considered as a good index of activation of the brain. It has been well established that c-fos and c-jun are activated during stress in broad areas of the brain. This study shows that the pigmentation positive rate and average absorbency of c-fos and c-jun in the right hippocampus and frontal lobe in the operation group are higher than those of the false operation group at different times after the operation (P > 0.05). In the operation group,the distinctions between the pigmentation positive rate and average absorbency of c-fos and c-jun at different times after the operation were significant,and increased with time after the operation (trend test P < 0.05). In the false operation group,the distinctions between the pigmentation positive rate and average absorbency of c-fos and c-jun at different times after the operation were insignificant (P > 0.05). This suggests that the expression of c-fos and c-jun has a negative correlation with the rCBF in the memory region.
The upregulation of c-jun and c-fos expression by cerebral ischemia may be due to the following factors: c-fos codes for a certain phosphoryl nucleoprotein, which acts as a transcription factor to activate the expression of target genes by binding to specific DNA sites; when new or intense stimulation is lacking,the transcription level of c-fos is low,but its expression is swiftly upregulated when the neuron is excited or its state is changed. Additionally,because the expression subsides within 5-6 h,c-fos immunohistochemistry can mark an excited neuron in multisynaptic neural pathways[9]. Thus,high level of c-fos detected by immunohistochemical methods can be regarded as the sign of changing neuronal activity,and it is widely applied in studies of functional nerve anatomy and neural pathways. Therefore,the decline of learning and memory function caused by cerebral ischemia may have a relationship with the enhanced expression of c-fos and c-jun in the hippocampus and frontal lobe caused by ischemia.
However,the mechanism of c-fos and c-jun promoting apoptosis has not been clarified yet,but presumably it is related with the effect of AP-1[10]. AP-1,which is activated by the expression of c-fos and c-jun (nucleus DNA-binding protein),is a heterodimer joined by a leucine zipper,and forms the structural base for transferring of apoptosis signals[11]. TPA response element (TRE) is present in its DNA binding region,and binds to DNA by each of its regions rich in alkalescent amino acid,regulating the replication and transcription of DNA. At the same time, it induces the expression of genes containing TRE, and acts as a gene regulating protein,regulating the transcriptions of target genes or the late onset genes, thereby,inducing apoptosis or promoting cell cycle. Alternatively,c-fos protein is regarded as a sort of membrane protein,which cross-links with antibodies to induce apoptosis. Therefore,the enhanced expression of c-fos and c-jun in the neurons of the regions involved in learning and memory can lead to the decline of learning and memory function.
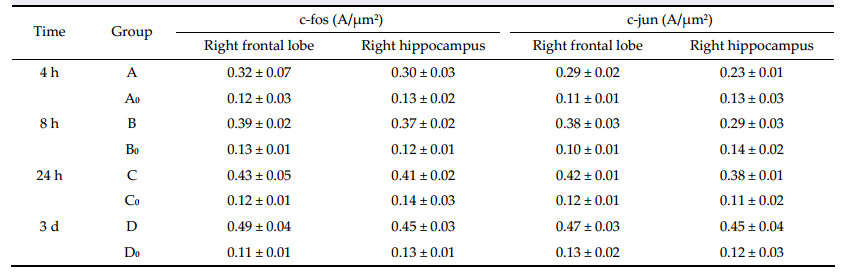
Study reports show that Bcl-2 and Bcl-xl proteins are anti-apoptotic,and Bax and Bcl-xs proteins promote apoptosis[12]. The excessive expression of Bcl-2 together with Bax forms isodimers,which inhibit apoptosis[13]. However,Bax-Bax isodimers can promote apoptosis, since Bax is an apoptosis promoting gene. It causes the releasing of some molecules,which leads to the activation of cysteine proteinase,and simultaneously, hastens apoptosis by resisting the protecting effect of Bcl-2 or induces apoptosis by conversion of mitochondrial transparency[14]. In this study,Bcl-2 and Bax positive cells are expressed in the operation group in the right hippocampus and frontal lobe at different time after the operation. The expression of Bcl-2,Bax, and the Bax/Bcl-2 protein expression ratio increased gradually (trend test P<0.05) with time after the operation. However,there was no expression of Bcl-2 and Bax positive cell in the false operation group at different time after the operation. This suggests that when the rCBF of the hippocampus and frontal lobe decreases,cerebral cell ischemia can lead to the increased expression of Bcl-2 and Bax positive cells. The enhanced Bax/Bcl-2 protein expression ratio promotes the cerebral cell apoptosis,which causes the decline of learning and memory function. Considering that the changes in the Bax/Bcl-2 ratio are linked to the survival of the cell and the increased Bax/Bcl-2 protein expression ratio can promote neuronal apoptosis,it is clear that the reduction of the rCBF can cause the elevation of the expression of Bcl-2,and Bax,and Bax/Bcl-2 ratio,which leads to decreased learning and memory function.
In conclusion,the mechanism underlying the learning and memory function decline caused by decreased rCBF is very complex. It not only involves the relationship of the above genes with apoptosis, but may also involve other causations,which are yet to be investigated. The next step should be studying that relationship between blood flow perfueion levels and other relevent gene of learning and memory function in strict control of various confounding factors,so improves the learning and memory function of patients with cerebral blood perfusion deficiency by controlling gene expression.
Acknowledgment We are grateful to M Yang,M Huang and M Wei for their technical assistance. Conflict of interests The authors have no financial interest to disclose regarding the article.| [1] | Cao YG, Xu LJ, Xu L, Song ZK, Wu JM. Progress in the mechanism and prevention of skeletal muscle ischemiareperfusion injury. Med Recapitul 2009, 15(1): 126-129. (in Chinese) |
| Click to display the text | |
| [2] | Séquier JM, Hunziker W, Richards G. Localization of calbindin D28 mRNA in rat tissues by in situ hybridization. Neurosci Lett 1988, 86(2): 155-160. |
| Click to display the text | |
| [3] | Ryseck RP, Bravo R. c-JUN, JUN B, and JUN D differ in their binding affinities to AP-1 and CRE consensus sequences: Effect of FOS proteins. Oncogene 1991, 6(4): 533-542. |
| Click to display the text | |
| [4] | Ohkubo Y, Arima M, Arguni E, Okada S, Yamashita K, Asari S, Obata S, Sakamoto A, Hatano M, O-Wang J, et al. A role for c-fos/Activator protein 1 in B lymphocyte terminal differentiation. J Immunol 2005, 174(12): 7703-7710. |
| Click to display the text | |
| [5] | Han J, Guo HZ. Effect of ischemic preconditioning on expression of hsp70 and fos after focal cerebral infarction in rat. Chin J Clin Neurosci 2004, 12(2): 136-139. (in Chinese) |
| Click to display the text | |
| [6] | Wiedmann R, Rosahl SK, Brinker T, Samii M, Nakamura M. Effect of acute and chronic bilateral visual deafferentation on c-fos immunoreactivity in the visual system of adult rats. Exp Brain Res 2013, 229(4): 595-607 |
| Click to display the text | |
| [7] | Montag-Sallaz M, Buonviso N. Altered odor-induced expression of c-fos and arg 3.1 immediate early genes in the olfactory system after familiarization with an odor. J Neurobiol 2002, 52(1): 61-72. |
| Click to display the text | |
| [8] | Zhou F, Guo JC, Yang R, Gu J, Jin HB, Wu GC, Cheng JS. Effects of taurine on cerebral blood flow perfusion, cell apoptosis, and infarct volume in acute cerebral ischemic rats. In Taurine 6. Oja SS, Saransaari P, Eds. US: Springer 2006, 583: 353-358. |
| Click to display the text | |
| [9] | Blasiak A, Siwiec M, Grabowiecka A, Blasiak T, Czerw A, Blasiak E, Kania A, Rajfur Z, Lewandowski MH, Gundlach AL. Excitatory orexinergic innervation of rat nucleus incertuse—Implications for ascending arousal, motivation and feeding control. Neuropharmacology 2015, 99: 432-447. |
| Click to display the text | |
| [10] | Herrera DG, Robertson HA. Activation of c-fos in the brain. Progr Neurobiol 1996, 50(2-3): 83-107. |
| Click to display the text | |
| [11] | Herdegen T, Waetzig V. AP-1 proteins in the adult brain: Facts and fiction about effectors of neuroprotection and neurodegeneration. Oncogene 2001, 20(19): 2424-2437. |
| Click to display the text | |
| [12] | Runyan JD, Dash PK. Distinct prefrontal molecular mechanisms for information storage lasting seconds versus minutes. Learn Mem 2005, 12(3): 232-238. |
| Click to display the text | |
| [13] | Hu JL, Xie H, Long ZJ, Meng Q, Zhang YH, He YS. Effects of Ning-Shen Wen-Dan decoction to the expression of apoptosis protein Bax, Bcl-2 in hippocampus of rats. Chin J Basic Med Tradit Chin Med 2013, 19(4): 394-396. (in Chinese) |
| Click to display the text | |
| [14] | Van Der Borght K, Wallinga AE, Luiten PGM, Eggen BJL, Van Der Zee EA. Morris water maze learning in two rat strains increases the expression of the polysialylated form of the neural cell adhesion molecule in the dentate gyrus but has no effect on hippocampal neurogenesis. Behav Neurosci 2005, 119(4): 926-932. |
| Click to display the text |