| Original Articles |
|


|
|
|
| Preliminary analysis of cellular sociology of co-cultured glioma initiating cells and macrophages in vitro |
| Mingxia Zhang1, Junjie Chen2, Lin Wang1, Xiaoyan Ji1, Lin Yang1, Yujing Sheng1, Hairui Liu1, Haiyang Wang1, Aidong Wang1, Xingliang Dai1, Xiaonan Li3, Qiang Huang1, Jun Dong1 |
1 Department of Neurology, the Second Affiliated Hospital of Soochow University, Suzhou 215004, China;
2 Department of Neurology, Xishan People's Hospital, Wuxi 214000, China;
3 Laboratory of Molecular Neuron-oncology, Texas Children's Cancer Center, Baylor College of Medicine, Houston, TX, USA |
|
|
|
|
Abstract Objective: Real-time monitoring of cytokine secretion at the single immunocyte level, based on the concept of immune cells, sociology has been recently reported. However, the relationships between glioma-initiating cells (GICs) and host immune cells and their mutual interactions in the tumor microenvironment have not been directly observed and remain unclear.
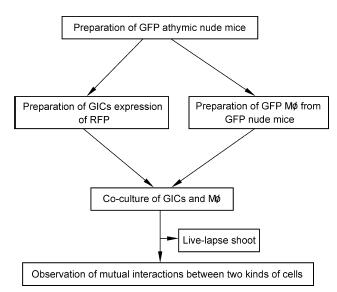
Methods: The dual fluorescence tracing technique was applied to label the co-cultured GICs and host macrophages (Mø), and the interactions between the two types of cells were observed using a live cell imaging system. Fusion cells in the co-culture system were monocloned and proliferated in vitro and their social interactions were observed and recorded.
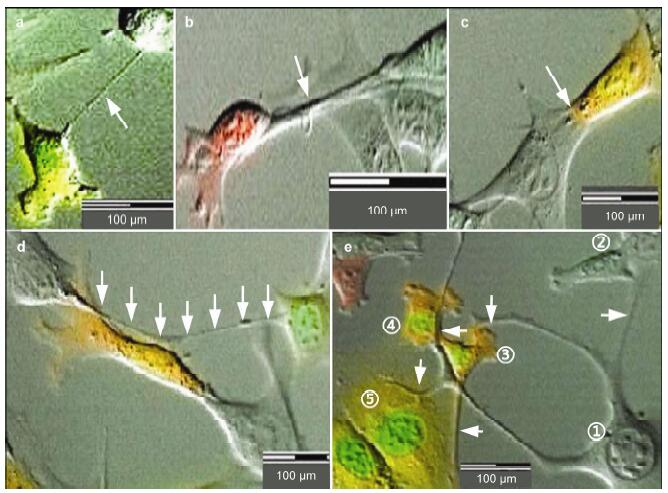
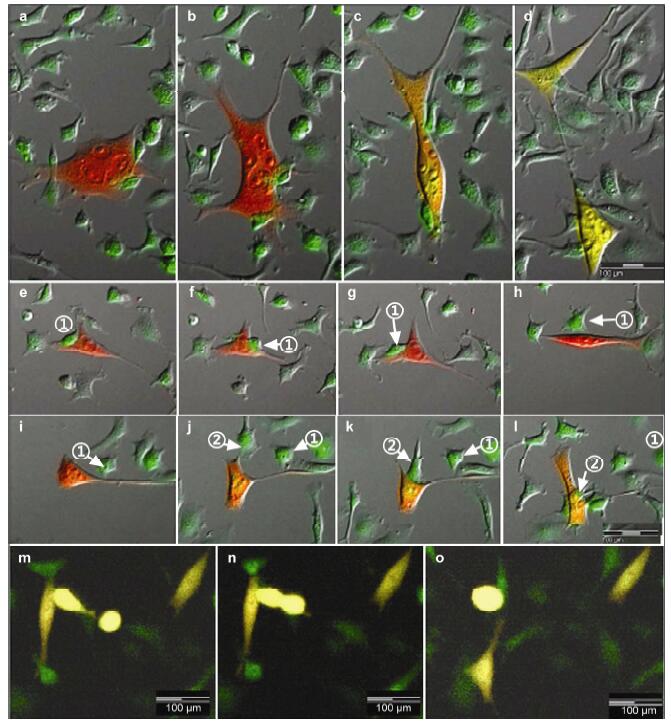
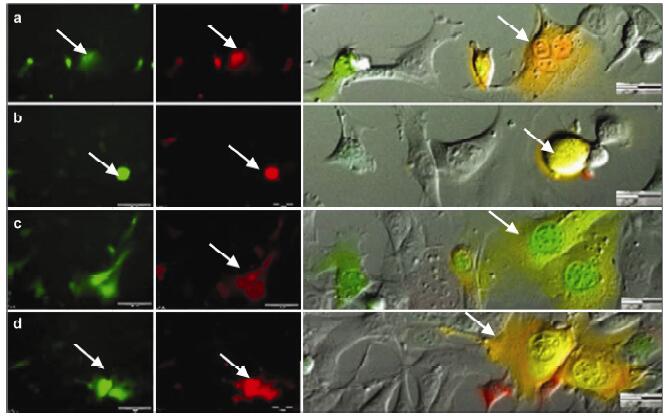
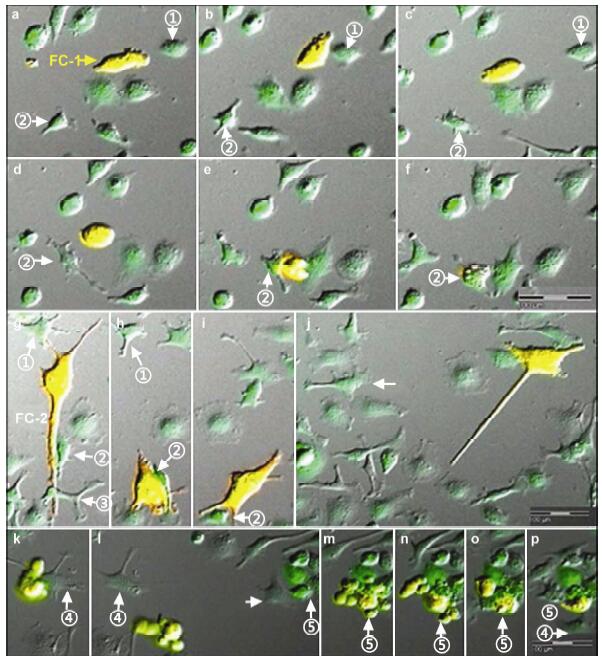
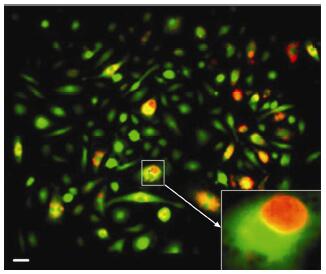
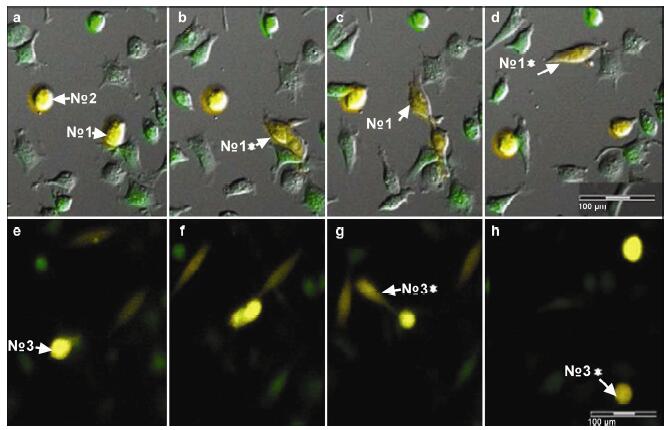
Results: Using real-time dynamic observation of target cells, 6 types of intercellular conjunction microtubes were found to function in the transfer of intercellular information between GICs and Mø; GICs and host Mø can fuse into hybrid cells after several rounds of mutual interactions, and then these fusion cells fused with each other; Fusion cells generated offspring cells through symmetrical and asymmetrical division or underwent apoptosis. A "cell in cell" phenomenon was observed in the fusion cells, which was often followed by cell release, namely entosis.
Conclusions: Preliminary studies revealed the patterns of cell conjunction via microtubes between GICs and host Mø and the processes of cell fusion, division, and entosis. The results revealed malignant transformation of host Mø, induced by GICs, suggesting complex social relationships among tumor-immune cells in gliomas.
|
|
Received: 20 May 2016
Published: 30 June 2016
|
| Fund: Supported by the National Natural Science Foundation of China (Grant No. 81472739) and the Natural Science Foundation of Jiangsu Province (Grant No. BK20151214). |
|
Corresponding Authors:
Jun Dong,E-mail:djdongjun@163.com
E-mail: djdongjun@163.com
|
|
Cite this article:
Mingxia Zhang, Junjie Chen, Lin Wang, Xiaoyan Ji, Lin Yang, Yujing Sheng, Hairui Liu, Haiyang Wang, Aidong Wang, Xingliang Dai, Xiaonan Li, Qiang Huang, Jun Dong. Preliminary analysis of cellular sociology of co-cultured glioma initiating cells and macrophages in vitro. Translational Neuroscience and Clinics, 2016, 2(2): 77-86.
URL:
http://tnc.tsinghuajournals.com/10.18679/CN11-6030/R.2016.018 OR http://tnc.tsinghuajournals.com/Y2016/V2/I2/77
|
|
|
[1] Wang AD, Dai XL, Cui BQ, Fei XF, Chen YM, Zhang JS, Zhang QB, Zhao YD, Wang ZM, Chen H, et al. Experimental research of host macrophage canceration induced by glioma stem progenitor cells. Mol Med Rep 2015, 11(4):2435-2442.
[2] Charles NA, Holland EC, Gilbertson R, Glass R, Kettenmann H. The brain tumor microenvironment. Glia 2012, 60(3):502-514.
[3] Wan Y, Fei XF, Wang ZM, Jiang DY, Chen HC, Yang J, Shi L, Huang Q. Expression of miR-125b in the new highly invasive glioma stem cell and progenitor cell line SU3. Chin J Cancer 2012, 31(4):207-214.
[4] Shirasaki Y, Yamagishi M, Shimura N, Hijikata A, Ohara O. Toward an understanding of immune cell sociology:Real-time monitoring of cytokine secretion at the single-cell level. IUBMB Life 2013, 65(1):28-34.
[5] Dong J, Zhang QB, Huang Q, Chen H, Shen YT, Fei XF, Zhang TY, Diao Y, Wu ZC, Qin ZH, et al. Glioma stem cells involved in tumor tissue remodeling in a xenograft model. J Neurosurg 2010, 113(2):249-260.
[6] Dong J, Zhao YD, Huang Q, Fei XF, Diao Y, Shen YT, Xiao H, Zhang TY, Lan Q, Gu XS. Glioma stem/progenitor cells contribute to neovascularization via transdifferentiation. Stem Cell Rev 2011, 7(1):141-152.
[7] Chen YM, Fei XF, Wang AD, Dai XL, Zhang JS, Cui BQ, Zhang QB, Zhao YD, Chen H, Wang ZM, et al. Host glial cell canceration induced by glioma stem cells in GFP/RFP dual fluorescence orthotopic glioma models in nude mice. Chin J Oncol 2013, 35(1):5-10.
[8] Hurtig J, Chiu DT, Önfelt B. Intercellular nanotubes:Insights from imaging studies and beyond. Wiley Interdiscip Rev Nanomed Nanobiotechnol 2010, 2(3):260-276.
[9] Osswald M, Jung E, Sahm F, Solecki G, Venkataramani V, Blaes J, Weil S, Horstmann H, Wiestler B, Syed M, et al. Brain tumour cells interconnect to a functional and resistant network. Nature 2015, 528(7580):93-98.
[10] Antonyak MA, Li B, Boroughs LK, Johnson JL, Druso JE, Bryant KL, Holowka DA, Cerione RA. Cancer cell-derived microvesicles induce transformation by transferring tissue transglutaminase and fibronectin to recipient cells. Proc Natl Acad Sci USA 2011, 108(12):4852-4857.
[11] Duelli D, Lazebnik Y. Cell fusion:A hidden enemy? Cancer Cell 2003, 3(5):445-448.
[12] Goldenberg DM, Zagzag D, Heselmeyer-Haddad KM, Berroa Garcia LY, Ried T, Loo M, Chang CH, Gold DV. Horizontal transmission and retention of malignancy, as well as functional human genes, after spontaneous fusion of human glioblastoma and hamster host cells in vivo. Int J Cancer 2012, 131(1):49-58.
[13] Schichor C, Albrecht V, Korte B, Buchner A, Riesenberg R, Mysliwietz J, Paron I, Motaln H, Turnšek TL, Jürchott K, et al. Mesenchymal stem cells and glioma cells form a structural as well as a functional syncytium in vitro. Exp Neurol 2012, 234(1):208-219.
[14] Chen EH, Olson EN. Unveiling the mechanisms of cell-cell fusion. Science 2005, 308(5720):369-373.
[15] Vignery A. Macrophage fusion:The making of osteoclasts and giant cells. J Exp Med 2005, 202(3):337-340.
[16] Vignery A, Gilgenkrantz S. Macrophage fusion:Are somatic and cancer cells possible partners? Med Sci (Paris) 2005, 21(12):1070-1075.
[17] Pawelek JM. Tumour cell hybridization and metastasis revisited. Melanoma Res 2000, 10(6):507-514.
[18] Pawelek JM, Chakraborty AK. Fusion of tumour cells with bone marrow-derived cells:A unifying explanation for metastasis. Nat Rev Cancer 2008, 8(5):377-386.
[19] Pawelek JM. Tumour-cell fusion as a source of myeloid traits in cancer. Lancet Oncol 2005, 6(12):988-993.
[20] Pawelek J, Chakraborty A, Lazova R, Yilmaz Y, Cooper D, Brash D, Handerson T. Co-opting macrophage traits in cancer progression:A consequence of tumor cell fusion? Contrib Microbiol 2006, 13:138-155.
[21] Duelli D, Lazebnik Y. Cell-to-cell fusion as a link between viruses and cancer. Nat Rev Cancer 2007, 7(12):968-976.
[22] Holmgren L, Bergsmedh A, Spetz AL. Horizontal transfer of DNA by the uptake of apoptotic bodies. Vox Sang 2002, 83(Suppl. 1):305-306.
[23] Dittmar T, Nagler C, Schwitalla S, Reith G, Niggemann B, Zänker KS. Recurrence cancer stem cells-made by cell fusion? Med Hypotheses 2009, 73(4):542-547.
[24] White E. Entosis:It's a cell-eat-cell world. Cell 2007, 131(5):840-842.
[25] Overholtzer M, Mailleux AA, Mouneimne G, Normand G, Schnitt SJ, King RW, Cibas ES, Brugge JS. A nonapoptotic cell death process, entosis, that occurs by cell-in-cell invasion. Cell 2007, 131(5):966-979. |
|
Viewed |
|
|
|
Full text
|
|
|
|
|
Abstract
|
|
|
|
|
Cited |
|
|
|
|
| |
Shared |
|
|
|
|
| |
Discussed |
|
|
|
|