摘要 There were two stages in the history of the studies on ascending reticular activating system of the brain (ARAS). The first stage began with the ARAS discovery by Magoun and Moruzzi and the following investigations using the methods of stimulation and lesion at that time mainly in acute cats. These studies led to the hypothesis of a "diffuse" and "unspecific" ARAS of the brain stem. The second stage was associated with using more precise neurophysiological and histochemical methods mainly in chronically operated free-moving cats and rats. By 2010, the idea of the ARAS as an organized hierarchy of the cerebral "waking centers" distributed along the entire cerebral axis and releasing all the known neuromediators of low molecular weight together with the most important neuropeptides was formulated. To date, the aforementioned hypothesis has been revised again. The glutamatergic activating system has been discovered and described in detail. Presumably, this system is responsible for the appearance of electroencephalogram (EEG) arousal reaction and maintenance of the neocortex in the state of tonic depolarization during wakefulness and rapid eye movement (REM) sleep. Its destruction results in a deep comatose-like state. At the same time, the activity of all other "waking centers" is probably the result of the cortical activation.
Abstract: There were two stages in the history of the studies on ascending reticular activating system of the brain (ARAS). The first stage began with the ARAS discovery by Magoun and Moruzzi and the following investigations using the methods of stimulation and lesion at that time mainly in acute cats. These studies led to the hypothesis of a "diffuse" and "unspecific" ARAS of the brain stem. The second stage was associated with using more precise neurophysiological and histochemical methods mainly in chronically operated free-moving cats and rats. By 2010, the idea of the ARAS as an organized hierarchy of the cerebral "waking centers" distributed along the entire cerebral axis and releasing all the known neuromediators of low molecular weight together with the most important neuropeptides was formulated. To date, the aforementioned hypothesis has been revised again. The glutamatergic activating system has been discovered and described in detail. Presumably, this system is responsible for the appearance of electroencephalogram (EEG) arousal reaction and maintenance of the neocortex in the state of tonic depolarization during wakefulness and rapid eye movement (REM) sleep. Its destruction results in a deep comatose-like state. At the same time, the activity of all other "waking centers" is probably the result of the cortical activation.
通讯作者:
Vladimir M.Kovalzon,E-mail:kovalzon@sevin.ru
E-mail: kovalzon@sevin.ru
引用本文:
Vladimir M. Kovalzon. Ascending reticular activating system of the brain[J]. 临床转化神经科学, 2016, 2(4): 275-285.
Vladimir M. Kovalzon. Ascending reticular activating system of the brain. Translational Neuroscience and Clinics, 2016, 2(4): 275-285.
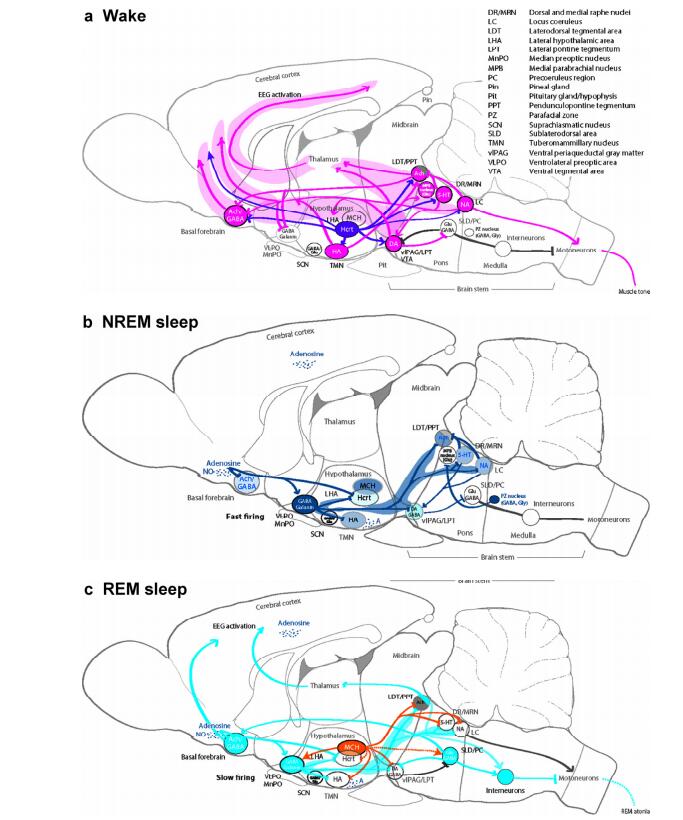
20170308103112 Figure 1 Sleep-wake regulation. (a) Wake: The brain stem arousal nuclei (pink) containing ACh, DA, 5-HT, or NA activate the thalamus, hypothalamus, spinal cord motor neurons, and the basal forebrain, and inhibit the VLPA (GABA, galanin); the hypothalamic arousal centers [pink: HA; dark purple: Hcrt] activate the cortex and arousal-related regions in the basal forebrain and brain stem; the thalamus activates the cortex. The glutamatergic MPB in the dorsal pontine tegmentum regulates arousal[28]. (b) NREM sleep: The hypothalamic preoptic area nuclei (dark blue), containing GABA and galanin, inhibit the brain stem and hypothalamic arousal nuclei; the endogenous sleep regulatory substances [adenosine and NO] inhibit basal forebrain arousal nuclei, hypocretin neurons, and TMN neurons; and adenosine activates VLPO neurons. NREM–REM switch: During NREM sleep, serotonergic DR and noradrenergic LC neurons inhibit LDT/PPT neurons. During NREM sleep, vlPAG/LPT neurons inhibit SLD/PC neurons. The GABAergic PZ in the pontomedullary junction promotes sleep[36]. (c) REM sleep: The REM-active brain stem nuclei, including LDT/PPT/SLD/PC and containing ACh, Glu, or GABA, promote activity in the basal forebrain and cortex and induce muscle atonia and rapid eye movements; hypothalamic neurons, containing MCH, promote REM sleep by suppressing REM-inhibitory brain centers, including vlPAG/LPT/DR/LC. NREM–REM switch: During REM sleep, DR/LC neurons become silent, enabling the cholinergic LDT/PPT neurons to generate the hallmarks of REM sleep, including rapid eye movements, EEG activation, and muscle atonia. This reciprocal activity between REM-on (LDT/PPT) and REM-off (DR/LC) neurons drives the cycling between REM and NREM sleep episodes. Additionally, GABAergic neurons participate in the mutual inhibition of REM-activating and REM-suppressing neurons. During REM sleep, SLD/PC neurons use ascending and descending projections to activate the cortex and promote muscle atonia.
[1]
Berger H. Üeber das elektroenkephalogramm des menschen: IL Mitteilung. J Psychol Neurol 1930, 40: 160-179.
[2]
Pelayo R,Dement WC. History of sleep physiology and medicine. In Principles and Practice of Sleep Medicine, 6th ed. Kryger M, Roth T, Dement WC, Eds. Philadelphia, PA: Elsevier, 2016, pp 3-14.
[3]
Moruzzi G, Magoun H. Brain stem reticular formation and activation of the EEG. Electroencephalogr Clin Neurophysiol 1949, 1(4): 455-473.
[4]
Koval'zon VM. Central mechanisms of the sleep-wakefulness cycle control. Hum Physiol 2011, 37(4): 500-508.
[5]
Brown RE, Basheer R, McKenna JT, Strecker RE, McCarley RW. Control of sleep and wakefulness. Physiol Rev 2012, 92(3): 1087-1187.
[6]
Lim MM, Szymusiak R. Neurobiology of arousal and sleep: updates and insights into neurological disorders. Curr Sleep Med Rep 2015, 1(2): 91-100.
[7]
Aston-Jones G, Chen S, Zhu Y, Oshinsky ML. A neural circuit for circadian regulation of arousal. Nat Neurosci 2001, 4(7): 732-738.
[8]
Steriade M, Timofeev I, Grenier F. Natural waking and sleep states: a view from inside neocortical neurons. J Neurophysiol 2001, 85(5): 1969-1985.
[9]
Steriade M, Timofeev I. Neuronal plasticity in thalamocortical networks during sleep and waking oscillations. Neuron 2003, 37(4): 563-576.
[10]
Timofeev I, Grenier F, Steriade M. Disfacilitation and active inhibition in the neocortex during the natural sleep-wake cycle: An intracellular study. Proc Natl Acad Sci USA 2001, 98(4): 1924-1929.
[11]
Timofeev I, Bazhenov M, Seigneur J, Sejnowski T. Neuronal synchronization and thalamocortical rhythms duringsleep, wake, and epilepsy. InJasper's Basic Mechanisms of the Epilepsies. 4th edn. Noebels JL, Avoli M, Rogawski MA, Olsen RW, Delgado-Escueta AV, Eds. New York: Oxford University Press, 2012, p 157.
[12]
Richter C, Woods IG, Schier AF. Neuropeptidergic control of sleep and wakefulness. Annu Rev Neurosci 2014, 37: 503-531.
Siegel JM. Rapid eye movement sleep. InPrinciples and Practice of Sleep Medicine, 6th ed. Kryger M, Roth T, Dement WC, Eds. Philadelphia PA: Elsevier, 2016, pp 78-95.
[15]
Koval'zon VM. The role of histaminergic system of the brain in the regulation of sleep-wakefulness cycle. Hum Physiol 2013, 39(6): 574-583.
[16]
Koval'zon VM. Brain and sleep: from neurons -to the molecules. Zh Vyssh Nerv Deiat Im I P Pavlova2013, 63(1): 48-60.
[17]
Kovalzon VM, Zavalko IM. The neurochemistry of the sleep-wakefulness cycle and Parkinson's disease. J Neurochem 2013, 7(3): 171-183.
[18]
Blanco-Centurion C, Gerashchenko D, Shiromani PJ. Effects of saporin-induced lesions of three arousal populations on daily levels of sleep and wake. J Neurosci 2007, 27(51): 14041-14048.
[19]
Denoyer M, Sallanon M, Buda C, Kitahama K, Jouvet M. Neurotoxic lesion of the mesencephalic reticular formation and/or the posterior hypothalamus does not alter waking in the cat. Brain Res 1991, 539(2): 287-303.
[20]
Jouvet M. Le sommeil, la conscience et l'éveil. Paris: Odile Jacob, 2016, p 216.
[21]
Carter ME, Yizhar O, Chikahisa S, Nguyen H, Adamantidis A, Nishino S, Deisseroth K, de Lecea L. Tuning arousal with optogenetic modulation of locus coeruleus neurons. Nat Neurosci 2010, 13(12): 1526-1533.
[22]
Kaur S, Junek A, Black MA, Semba K. Effects of ibotenate and <sup>192</sup>IgG-saporin lesions of the nucleus basalis magnocellularis/ substantia innominata on spontaneous sleep and wake states and on recovery sleep after sleep deprivation in rats. J Neurosci 2008, 28(2): 491-504.
[23]
Gerashchenko D, Chou TC, Blanco-Centurion CA, Saper CB, Shiromani PJ. Effects of lesions of the histaminergic tuberomammillary nucleus on spontaneous sleep in rats. Sleep 2004, 27(7): 1275-1281.
[24]
Lu J, Jhou TC, Saper CB. Identification of wake-active dopaminergic neurons in the ventral periaqueductal gray matter. J Neurosci 2006, 26(1): 193-202.
[25]
Shouse MN, Siegel JM. Pontine regulation of REM sleep components in cats: integrity of the pedunculopontine tegmentum (PPT) is important for phasic events but unnecessary for atonia during REM sleep. Brain Res 1992, 571(1): 50-63.
[26]
Webster HH, Jones BE. Neurotoxic lesions of the dorsolateralpontomesencephalic tegmentum-cholinergic cell area in the cat. Ⅱ. Effectsupon sleep-waking states. Brain Res 1988, 458(2): 285-302.
[27]
Saper CB, Fuller PM, Pedersen NP, Lu J, Scannell TE. Sleep state switching. Neuron 2010, 68(6): 1023-1042.
[28]
Fuller P, Sherman D, Pedersen NP, Saper CB, Lu J. Reassessment of the structural basis of the ascending arousal system. J Comp Neurol 2011, 519(5): 933-956.
[29]
Villablanca JR. Counterpointing the functional role of the forebrain and of thebrainstem in the control of the sleep-waking system. J Sleep Res 2004, 13(3): 179-208.
[30]
Steriade M, McCarley RW. Brainstem Control of Wakefulness and Sleep. US: Springer, 1990.
[31]
Xu M, Chung S, Zhang SY, Zhong P, Ma CY, Chang WC, Weissbourd B, Sakai N, Luo LQ, Nishino S. Basal forebrain circuit for sleep-wake control. Nat Neurosci 2015, 18(11): 1641-1647.
[32]
Kaur S, Pedersen NP, Yokota S, Hur EE, Fuller PM, Lazarus M, Chamberlin NL, Saper CB. Glutamatergic signaling from the parabrachial nucleus plays a critical role in hypercapnic arousal. J Neurosci 2013, 33(18): 7627-7640.
[33]
Peng B, Zhang S, Dong H, Lu Z. Clinical, histopathological and genetic studies in a case of fatal familial insomnia with review of the literature. Int J Clin Exp Pathol 2015, 8(9): 10171-10177.
[34]
Haas HL, Lin JS. Waking with the hypothalamus. Pflugers Arch Eur J Physiol 2012, 463(1): 31-42.
[35]
Anaclet C, Ferrari L, Arrigoni E, Bass CE, Saper CB, Lu J, Fuller PM. The GABAergic parafacial zone is a medullary slow wave sleep-promoting center. Nat Neurosci 2014, 17(9): 1217-1224.
[36]
Anaclet C, Lin JS, Vetrivelan R, Krenzer M, Vong L, Fuller PM, Lu J. Identification and characterization of a sleep-active cell group in the rostral medullary brainstem. J Neurosci 2012, 32(50): 17970-17976.
[37]
Reinoso-Suárez F, De Andrés I, Rodrigo-Angulo LM, Rodríguez-Veiga E. Location and anatomical connections of a paradoxical sleep induction site in the cat ventral pontine tegmentum. Eur J Neurosci 1994, 6(12): 1829-1836.
[38]
Weber F, Chung S, Beier KT, Xu M, Luo LQ, Dan Y. Control of REM sleep by ventral medulla GABAergic neurons. Nature 2015, 526(7573): 435-438.
[39]
Luppi PH, Clément O, Fort P. Paradoxical (REM) sleep genesis by the brainstem is under hypothalamic control. Curr Opin Neurobiol 2013, 23(5): 786-792.
 2016, Vol. 2
2016, Vol. 2